I recorded the TV show Evolve:Eyes, which aired on the History Channel last night, and I watched it today. Last night during the original airing, I was in LA watching the Dodgers with my family. I think my son was quite taken with professional baseball. This was not his first game (I had to make sure that for both kids, their first game was in Milwaukee, where mine was), but I don’t think my son remembers seeing the Brewers in Miller Park. So when we walked through the gate of Dodger Stadium, I saw the awe in his eyes, as he took in the stadium’s brightness, the impossibly green grass, and as we heard 40,000 fans cheer together. There is something to be said for appearance and spectacle, which can make even things like playing baseball games seem important.
This was the primary strength that I found in the History Channel show Evolve:Eyes; it looked really slick; it had its share of spectacle. There were fantastic graphics. Three dimensional eye figures exploding into component lens, cornea, and retina. The linear progression of eye evolution was re-told graphically, a few photoreceptors morphing into a deepening pit. Then a lens appeared, until the human eye had “evolved” right before my own eyes.
There were also dramatic film sequences of real scientists. There was a re-enactment of my friend (we were postdocs together in Chicago) Nate Dominy of UCSC doing his field research observing primates (I’m guessing they filmed Nate in Santa Cruz with binoculars and cut in footage of primates). There was footage of my friend Chris Kirk (haven’t seen him for a while, but we were grad students at Duke at the same time) pulling eye balls out of jars, complete with lab coat (most biologists I know don’t wear lab coats, unless they’re being filmed, or maybe if their pulling eyeballs out of a smelly formalin/ethanol solution). And there was footage of my colleague Alex Goodell collecting Polyorchus (a cnidarian) and shining LED lights on them to watch their behavior: green light, then blue, illuminating the tank and animal like neon lights in the dark. Scott Edwards, alone in an arid landscape, artistically filmed full-body and off-center, commented on how many times eyes evolved, my proverbial red herring, deciding to re-tell the take of a fellow Harvard-man, the late Ernst Mayr. Spectacle. Beautiful.
But all was not positive from my perspective. Namely, the show fell into the two most common traps that plague the understanding of evolution by the public. 1) Evolution is not adaptive story telling and 2) Evolution has not progressed in a direct line from simple to complex; evolution most certainly does not culminate with the human species. In nature, humans are no more special than bacteria. We are just another twig on the great tree of life (that was for Eisen).
Very little of this show was actually about evolution, and most of the “evolution” that was mentioned did not explain the actual science behind it (there were of course some excpetions). Instead, there was a lot of adaptive storytelling. Eagles evolved amazing vision to allow them to be better hunters. I don’t doubt this is true, but this show very often simply asserted and explained current function, with little mention of how evolution of those traits proceeded. Jellyfish try to escape UV/blue because it is damaging. Again, this may be true, but it was simply asserted in the show. Evolution was simply assumed, with little mention of how we actually have established that evolution works.
But the most egregious to me was how the narrative and graphics re-enforced the non-existent linear progression from jellyfish to humans. In fact, the graphics were a line, shown between stories, proceeding from 600 million years ago to present day primates. In this hypothetical line, first came the “jellyfish” (they actually showed a hydromedusan). The camera simulated the “primitive” jelly’s blurry vision, far inferior to our own, no mention of the cubomedusan’s highly complex eye. Next came the trilobites, whose calcite compound eyes are among the oldest fossil eyes. (To the show’s credit, they did mention that trilobites are not direct ancestors of living arthropods, a slight nod to “tree thinking”. Also, the Cambrian and trilobite animations were pretty cool. More spectacle.)
Then came the vertebrates. Not much mention of fish, instead we jump to dinosaurs. More flash here, tremendous spectacle, shining lasers on models of T. rex, the icon of charismatic species. Then the story moves up the ladder to early mammals, “primitive mammals” eeking out an existence, with only “black and white” vision, living in the shadow of dinosaurs. Then the great KT event knocks out rex and mammals can finally flourish. This next bit really got me. They compare human (and some other primates’) vision to that of early mammals, saying how great we are to be able to see red. What a shallow point of comparison, just to inflate the human ego, and maintain the march of progress up to humans! Why not compare to our eyes to stomatopods (mantis shrimps), who see circular polarized light, and have upwards of 12-chromatic compared to our tri-chromatic vision. Or even compare us to adult maggots (i.e. flies), who see polarized and UV light that we cannot detect. In my book, the evolutionary ladder of eyes proceeds up to mantis shrimp, not humans.
But perhaps I am being a bit too critical, expecting too much nuance from a show aimed to keep viewers from switching to, say, ESPN to see how the Dodgers are doing (they won by the way). Spectacle can be something to celebrate!
After the Dodger’s game last night, I could tell my son was inspired by spectacle, not by nuance. In fact, I doubt he got any nuances of the game. He was oblivious to strategic pitching changes; clueless about a rare call that gave the Dodgers an extra run in a tight game (the Giant's left fielder juggled the ball out of play, something I’ve never seen, after many years of watching baseball, e.g. I’ve seen in person a triple play, and Ricky Henderson’s record breaking stolen base). No, it wasn’t nuance that compelled my son to ask when he gets to start soccer, and if I can video record his games so we can watch them on TV. It was spectacle, fan screaming, light flashing, ice cream in a Dodger helmet spectacle.
So who am I to complain if somewhere, some kid is taken by the spectacle of eye evolution, as so vividly portrayed last night? Taken by the idea of shining lasers on T. rex, or neon LED’s on pulsating jellyfish in dark rooms? Maybe an African-American kid saw Scott Edwards on TV last night, and dreamed a dream of studying evolution. Maybe the child of an anti-evolutionist now has some questions.
There is a place for spectacle. If spectacle can inspire, nuance can come later.
Wednesday, July 30, 2008
Thursday, July 24, 2008
Ostra-blog 1 - Gigantocypris
I study ostracods, or ostracodes, depending on your preference. Not octapods, as my father-in-law might tell you, but ostracods: Crustaceans of the class Ostracoda. Why you might ask? Well, the origin story is told at the beginning of this post.
Still, you may be asking, WHAT is an ostracod anyway? Many people, even biologists have never heard of an ostracod. This is why I carry a picture of an ostracods in my wallet, sandwiched between my credit cards and photos of my kids. Well, to promote ostracods to the seven readers of this blog, I've decided I would like to start a series of posts introducing ostracods and their amazing biology. I shall dub this series my "ostra-blogs". To kick things off, I'd like to tell you a little bit about Gigantocypris.
Gigantocypris is a deep-sea ostracod, living at around 1000m depth as plankton. Most ostracods are pretty small around half to 2 mm - barely visible to pepper corn sized. Gigantocypris, true to their name are ostracod giants, up to 3 cm in width - ping pong ball-sized. They are caught from big research ships in plankton nets. I've never had the pleasure of collecting one myself, but a colleague very kindly sent one to me by FedEx, so I could see one alive. In life, where people live at least, they are orange-red in color. But in their native deep sea habitat, long wavelength light does not penetrate the ocean very far, so their orange pigment would not reflect any light. Another kind colleague sent me some preserved specimens in ethanol from a ship that sailed the Pacific off South America. The alcohol preservative bleaches pigments and the ostracods turn a ghostly, semi-opaque white. When I grab the test tube, I always think of cocktail onions floating the the liquid - Gigantocypris martini anyone?
Perhaps the most amazing feature of Gigantocypris is their eyes. In most animals' eyes, light is bent and focused with lenses. But a select few bend light with mirrors. Gigantocypris is one of the few, bearing a huge pair of parabolic reflectors behind light sensitive patches. The naturalist Alistair Hardy described the animals this this way in 1959:
Mike Land worked out the optics of these incredible eyes. Gigantocypris doesn't focus light beams on to a single point. Instead, a line of focus is made along an elongated retina. These animals probably are maximizing the detection of light out to their sides. There is essentially no sunlight at the depths where we find Gigantocypris, so Land suggests that Gigantocypris is detecting light created by bioluminescent animals. Gigantocypris is, in fact, a predator. Small fishes have even been found in their guts, great fun for an ostracodologist like me, used to painstakingly sorting sand grains from ostracods!
Gigantocypris is probably the most famous of ostracods (with the possible exception of Umihotaru, which will be the subject of another ostra-blog, I'm sure). Gigantocypris having made it to MBARI's deep sea poster (mid-right, #26) and if you watch Blue Planet carefully, you can catch a video glimpse.
I will leave you with some fantastic photos of this beautiful animal that are available on the web.

Charismatic aren't they? (Picture from this polar scientist''s blog)

Other images are also online. Try this search.
Still, you may be asking, WHAT is an ostracod anyway? Many people, even biologists have never heard of an ostracod. This is why I carry a picture of an ostracods in my wallet, sandwiched between my credit cards and photos of my kids. Well, to promote ostracods to the seven readers of this blog, I've decided I would like to start a series of posts introducing ostracods and their amazing biology. I shall dub this series my "ostra-blogs". To kick things off, I'd like to tell you a little bit about Gigantocypris.
Gigantocypris is a deep-sea ostracod, living at around 1000m depth as plankton. Most ostracods are pretty small around half to 2 mm - barely visible to pepper corn sized. Gigantocypris, true to their name are ostracod giants, up to 3 cm in width - ping pong ball-sized. They are caught from big research ships in plankton nets. I've never had the pleasure of collecting one myself, but a colleague very kindly sent one to me by FedEx, so I could see one alive. In life, where people live at least, they are orange-red in color. But in their native deep sea habitat, long wavelength light does not penetrate the ocean very far, so their orange pigment would not reflect any light. Another kind colleague sent me some preserved specimens in ethanol from a ship that sailed the Pacific off South America. The alcohol preservative bleaches pigments and the ostracods turn a ghostly, semi-opaque white. When I grab the test tube, I always think of cocktail onions floating the the liquid - Gigantocypris martini anyone?
Perhaps the most amazing feature of Gigantocypris is their eyes. In most animals' eyes, light is bent and focused with lenses. But a select few bend light with mirrors. Gigantocypris is one of the few, bearing a huge pair of parabolic reflectors behind light sensitive patches. The naturalist Alistair Hardy described the animals this this way in 1959:
The paired eyes have huge metallic-looking reflectors
behind them, making them appear like the headlamps of
a large car; they look out through glass-like windows in
the otherwise orange carapace and no doubt these concave
mirrors behind serve instead of a lens in front.
Mike Land worked out the optics of these incredible eyes. Gigantocypris doesn't focus light beams on to a single point. Instead, a line of focus is made along an elongated retina. These animals probably are maximizing the detection of light out to their sides. There is essentially no sunlight at the depths where we find Gigantocypris, so Land suggests that Gigantocypris is detecting light created by bioluminescent animals. Gigantocypris is, in fact, a predator. Small fishes have even been found in their guts, great fun for an ostracodologist like me, used to painstakingly sorting sand grains from ostracods!
Gigantocypris is probably the most famous of ostracods (with the possible exception of Umihotaru, which will be the subject of another ostra-blog, I'm sure). Gigantocypris having made it to MBARI's deep sea poster (mid-right, #26) and if you watch Blue Planet carefully, you can catch a video glimpse.
I will leave you with some fantastic photos of this beautiful animal that are available on the web.

Charismatic aren't they? (Picture from this polar scientist''s blog)
The picture below is a side view. This one is "pregnant".
Other images are also online. Try this search.
Tuesday, July 22, 2008
Eye evolution: post-dinosaur, pre-Noah's Ark
Back in January or so, I was interviewed by researchers at the History Channel over the phone. They were working on a new series on evolution, and one episode was to be about eyes. I guess they had seen our lab's work last year on cnidarian opsins in the news and decided to talk to me about it. Looks like they might mention this and/or research by Suga and Gehring published in Janurary on cnidarian opsins. Here is the blurb from the History Channel website:
They are one of evolution's most useful and prevalent inventions. Ninety five percent of living species are equipped with eyes and they exist in many different forms. Learn how the ancestors of jellyfish may have been the first to evolve light-sensitive cells. Discover how dinosaur's evolved eyes that helped them become successful hunters. Finally, learn how primates evolved unique adaptations to their eyes that allowed them to better exploit their new habitat, and how the ability to see colors helped them find food.
I remember from the interview that they were really probing for flashy stuff. They wanted to know what they would see if they came to film at UCSB. They didn't seem too interested in filming hydra sitting in a dish (although that's one of the things we are doing in the lab this summer). I can see from the abstract that they settled on dinosaurs and primates - the proverbial charismatic megafauna....
I just looked up the schedule, and the eye evolution program will air just after Dinosaur Fight Club, and just before a documentary about a flood 3,000 years ago that they are dubbing Noah's flood! Should be fun!
The date is Tuesday July 29, eye evolution at 10 pm.
I'll try to give an opinion on the Eye Evolution program here after it airs.
They are one of evolution's most useful and prevalent inventions. Ninety five percent of living species are equipped with eyes and they exist in many different forms. Learn how the ancestors of jellyfish may have been the first to evolve light-sensitive cells. Discover how dinosaur's evolved eyes that helped them become successful hunters. Finally, learn how primates evolved unique adaptations to their eyes that allowed them to better exploit their new habitat, and how the ability to see colors helped them find food.
I remember from the interview that they were really probing for flashy stuff. They wanted to know what they would see if they came to film at UCSB. They didn't seem too interested in filming hydra sitting in a dish (although that's one of the things we are doing in the lab this summer). I can see from the abstract that they settled on dinosaurs and primates - the proverbial charismatic megafauna....
I just looked up the schedule, and the eye evolution program will air just after Dinosaur Fight Club, and just before a documentary about a flood 3,000 years ago that they are dubbing Noah's flood! Should be fun!
The date is Tuesday July 29, eye evolution at 10 pm.
I'll try to give an opinion on the Eye Evolution program here after it airs.
Tuesday, July 15, 2008
Evolutionary Origins of Phototransduction
Below is a section of an article I am drafting on eye evolution. Does it make sense? Any parts impossible to understand? Comments are welcomed!
In The Origin of Species, Darwin hypothesized that the first step in the evolution of eyes involved the gain of photosensitivity in a nerve, writing, “…I may remark that several facts make me suspect that any sensitive nerve may be rendered sensitive to light…” Here, Darwin was making the assumption that variation leading to light sensitivity of previously non-light-sensitive nerves was abundant, and that natural selection could then act on that variation. But how did those specific variations originate? How does a nerve become light sensitive? In his notebook, Darwin suggested that it might be impossible to understand how a nerve gained light sensitivity. He wrote “to show how the first eye is formed, — how one nerve becomes sensitive to light, ...is impossible.” Since the mechanisms of heredity and the molecular mechanisms of photoreception were unknown in Darwin’s time, no specific hypothesis could even be posed about the genesis of light perception. One hundred and fifty years later, things have changed.
Today, through an understanding of the distinct evolutionary histories of the components of vision, and more specifically of phototransduction (the signaling pathway that turns light into a nerve signal), we can now pose a specific historical hypothesis for their origin: Phototransduction originated within animals by modifying an existing signaling pathway. More specifically, at some time before the divergence of jellyfish and humans, but likely after thesplit common ancestor of sponges and humans, the first light sensitive animal opsin protein originated. This protein did not originate from nothing, nor was it newly breathed into an ancient animal genome by a designer. Instead, opsins arose by mutation of an existing receptor to render it light sensitive. This historical hypothesis makes numerous predictions, and available data are consistent with the hypothesis. It also makes predictions that have not yet been tested, indicating promising areas for future research.
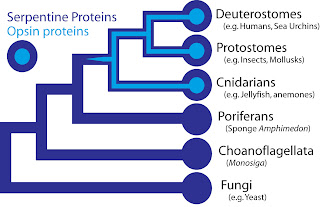
If the hypothesis that phototransduction arose within animals is valid, then some components of phototransduction should exist within animals, but should not exist (or should possess a different function) outside of animals. Such is the case for opsin, which is present as the primary photopigment gene in most animals. Recent research shows that various cnidarians, including a hydra, a sea anemone, a hydrozoan, and a box jellyfish, possess opsins (Plachetzki et al, 2007; Suga et al, 2008; Kozmik et al, 2008). At the same time, opsins are absent from sponges and non-animals (See Figure). In science, demonstrating the absence of something like a gene is difficult, because a skeptic can always invent a reason why the target was accidentally missed. In the case the opsins in question, scientists have determined the entire genome sequence of the sponge Amphimedon queenslandica, the choanoflagellate Monosiga and numerous fungi, making the presence of opsin in those organisms very unlikely. Some proteins of near-animals are in fact rather similar to opsins, but in every case the non-animal receptors lack characteristics that specifically define opsins. Therefore, although opsins might have been present at the origin of animals and lost in sponges, their absence instead strongly suggests that they originated within animals, before thesplit common ancestor of humans, insects, and cnidarians, all of which possess opsins.
With current knowledge that opsin is the basis of light sensitivity, Darwin’s question of how a nerve becomes light sensitive can be rephrased as, “how did animal opsins originate”? Proteins rarely originate from nowhere, and opsins are no exception. Opsins form a subfamily within a larger family of proteins called G-protein coupled receptors (GPCRs), also sometimes called serpentine proteins because they snake back and forth across cell membranes. Since serpentine proteins are present in all animals and their close relatives - including sponges, Monsiga, and fungi - this broad class of proteins long predates animals. In yeast (a fungus), these receptors are sensitive to pheromones and they even direct a signal through proteins homologous to non-opsin phototransduction proteins. As such, a signaling pathway exists outside animals, which is very similar to phototransduction, except the receptor protein detects pheromones, not light. Receptors outside animals share some characteristics with opsin, like snaking through a cell membrane seven times. It is one of these serpentine proteins that served as the progenitor of the first opsin protein, as evidenced by the similarity of opsins and other serpentine proteins.
Darwin’s question can be refined farther, to “how did a serpentine protein gain the ability to respond to light?” And since opsin’s light sensitivity is mediated by its ability to bind a light reactive chemical, called a chromophore, the question can be even further refined to ask how a GPCR must be modified to bind a chromophore. In the case of opsin, we know that a particular amino acid – a Lysine in the 7th membrane-spanning region – binds to the light reactive chemical. Presumably then, a mutation changing an amino acid in the 7th trans-membrane region of a light insensitive GPCR was involved in the acquisition of light sensitivity in animals. This fateful mutation coupled with numerous other mutations, are responsible for the origins of eyes and vision. Although science has not yet tracked down every single mutation involved in the evolution of vision, the origin of opsins clearly illustrate, in richer detail than Darwin might have imagined, the natural processes that gradually allow the evolution of complex features.
In The Origin of Species, Darwin hypothesized that the first step in the evolution of eyes involved the gain of photosensitivity in a nerve, writing, “…I may remark that several facts make me suspect that any sensitive nerve may be rendered sensitive to light…” Here, Darwin was making the assumption that variation leading to light sensitivity of previously non-light-sensitive nerves was abundant, and that natural selection could then act on that variation. But how did those specific variations originate? How does a nerve become light sensitive? In his notebook, Darwin suggested that it might be impossible to understand how a nerve gained light sensitivity. He wrote “to show how the first eye is formed, — how one nerve becomes sensitive to light, ...is impossible.” Since the mechanisms of heredity and the molecular mechanisms of photoreception were unknown in Darwin’s time, no specific hypothesis could even be posed about the genesis of light perception. One hundred and fifty years later, things have changed.
Today, through an understanding of the distinct evolutionary histories of the components of vision, and more specifically of phototransduction (the signaling pathway that turns light into a nerve signal), we can now pose a specific historical hypothesis for their origin: Phototransduction originated within animals by modifying an existing signaling pathway. More specifically, at some time before the divergence of jellyfish and humans, but likely after the

If the hypothesis that phototransduction arose within animals is valid, then some components of phototransduction should exist within animals, but should not exist (or should possess a different function) outside of animals. Such is the case for opsin, which is present as the primary photopigment gene in most animals. Recent research shows that various cnidarians, including a hydra, a sea anemone, a hydrozoan, and a box jellyfish, possess opsins (Plachetzki et al, 2007; Suga et al, 2008; Kozmik et al, 2008). At the same time, opsins are absent from sponges and non-animals (See Figure). In science, demonstrating the absence of something like a gene is difficult, because a skeptic can always invent a reason why the target was accidentally missed. In the case the opsins in question, scientists have determined the entire genome sequence of the sponge Amphimedon queenslandica, the choanoflagellate Monosiga and numerous fungi, making the presence of opsin in those organisms very unlikely. Some proteins of near-animals are in fact rather similar to opsins, but in every case the non-animal receptors lack characteristics that specifically define opsins. Therefore, although opsins might have been present at the origin of animals and lost in sponges, their absence instead strongly suggests that they originated within animals, before the
With current knowledge that opsin is the basis of light sensitivity, Darwin’s question of how a nerve becomes light sensitive can be rephrased as, “how did animal opsins originate”? Proteins rarely originate from nowhere, and opsins are no exception. Opsins form a subfamily within a larger family of proteins called G-protein coupled receptors (GPCRs), also sometimes called serpentine proteins because they snake back and forth across cell membranes. Since serpentine proteins are present in all animals and their close relatives - including sponges, Monsiga, and fungi - this broad class of proteins long predates animals. In yeast (a fungus), these receptors are sensitive to pheromones and they even direct a signal through proteins homologous to non-opsin phototransduction proteins. As such, a signaling pathway exists outside animals, which is very similar to phototransduction, except the receptor protein detects pheromones, not light. Receptors outside animals share some characteristics with opsin, like snaking through a cell membrane seven times. It is one of these serpentine proteins that served as the progenitor of the first opsin protein, as evidenced by the similarity of opsins and other serpentine proteins.
Darwin’s question can be refined farther, to “how did a serpentine protein gain the ability to respond to light?” And since opsin’s light sensitivity is mediated by its ability to bind a light reactive chemical, called a chromophore, the question can be even further refined to ask how a GPCR must be modified to bind a chromophore. In the case of opsin, we know that a particular amino acid – a Lysine in the 7th membrane-spanning region – binds to the light reactive chemical. Presumably then, a mutation changing an amino acid in the 7th trans-membrane region of a light insensitive GPCR was involved in the acquisition of light sensitivity in animals. This fateful mutation coupled with numerous other mutations, are responsible for the origins of eyes and vision. Although science has not yet tracked down every single mutation involved in the evolution of vision, the origin of opsins clearly illustrate, in richer detail than Darwin might have imagined, the natural processes that gradually allow the evolution of complex features.
Wednesday, July 9, 2008
Choose your own descent
I was interested to see that over at The Tree of Life Jonathan Eisen discussed a few different metaphors for common descent, which include:
The tree of life
The coral of life
The watershed of life
Blood vessels of life
Lungs of life
Shrub of life
I think metaphors are useful for teaching the idea of common descent and tree thinking, and so I try to think of new metaphors from time to time, since I teach a course on Macroevolution and also talk a lot about phylogenies in an Invertebrate Zoology course.
One that I think is really powerful, is found in Richard Dawkins' book Ancestor's Tale. The idea there is to reverse time in common descent, and imagine humans walking on a path to join our ancestors. First chimps join the band, later other apes, then other primates, other mammals, etc. This analogy is good for understanding the history of a particular entity, like the human species. However, since it has a specific starting point, it tends to re-enforce the way people conceive of evolution as a linear series of events. This is true if we have one specific endpoint in mind (like humans), but evolution does not have one specific endpoint.
So, this morning, this was going around in my head, and a new metaphor sprung into my brain: A choose your own adventure book. These books have a section of the story, and then the reader must choose between two (or more) different paths, each with a different outcome to the story.
Congratulations, your character has just achieved bilateral symmetry! If you would like to proceed down the path of protostomy, go to page 150. If you would like to proceed down the path of deuterostomy, go to page 376. It might be fun to write an actual evolution book this way!
The tree of life
The coral of life
The watershed of life
Blood vessels of life
Lungs of life
Shrub of life
I think metaphors are useful for teaching the idea of common descent and tree thinking, and so I try to think of new metaphors from time to time, since I teach a course on Macroevolution and also talk a lot about phylogenies in an Invertebrate Zoology course.
One that I think is really powerful, is found in Richard Dawkins' book Ancestor's Tale. The idea there is to reverse time in common descent, and imagine humans walking on a path to join our ancestors. First chimps join the band, later other apes, then other primates, other mammals, etc. This analogy is good for understanding the history of a particular entity, like the human species. However, since it has a specific starting point, it tends to re-enforce the way people conceive of evolution as a linear series of events. This is true if we have one specific endpoint in mind (like humans), but evolution does not have one specific endpoint.
So, this morning, this was going around in my head, and a new metaphor sprung into my brain: A choose your own adventure book. These books have a section of the story, and then the reader must choose between two (or more) different paths, each with a different outcome to the story.
Congratulations, your character has just achieved bilateral symmetry! If you would like to proceed down the path of protostomy, go to page 150. If you would like to proceed down the path of deuterostomy, go to page 376. It might be fun to write an actual evolution book this way!
Monday, July 7, 2008
Box Jellies and the Red Herring of Eye Evolution
Smoking a herring turns it red and imbues the fishy flesh with an extra strong odor, which can be used to throw bloodhounds off your trail, or to train hunting dogs to stay on the main scent trail, even if far less odoriferous. Such are a few hypotheses for the origin of the term "red herring", used to denote an enticing, yet ultimately uninformative beacon.
I've come to realize that my own field of eye evolution has a resoundingly pungent red herring that continually throws scientists and non-scientists alike off the trail of a deep understanding of how eyes, and more generally, photoreception, evolved. In this case, the rank fish is the simple and seemingly harmless question, "How many times did eyes evolve?"
Quite often, when I tell a newly acquainted colleague that I study eye evolution, he or she will pull the red herring out of their back pocket, and dash across the room, dragging the fish behind them. "So how many times did eyes evolve, anyway?” they ask, hoping I have a key to this deep mystery. When I was a younger bloodhound, I often took that bait, howling and bounding enthusiastically after that trail, circling the room again and again. But I've followed that trail enough times by now to know that it circles back on itself in a confusing and illogical way.
How many times eyes evolved, though enticingly simple, is an all but unanswerable question; at least in terms of a numerical answer that people crave. Eyes did not evolve one time, nor did they evolve forty to sixty-five times. The answer is that some components of animal eyes are shared in all animal eyes we've ever examined. Yet some components are new comers to organs that probably functioned as eyes even before those components joined the party. Asking how many times eyes evolved makes the implicit assumption that every component of that eye shares a congruent history, that an eye is either all there, or all not there. But eyes are complicated structures, and the evolutionary histories of its parts are varied.
A new paper by Kozmik et al in PNAS is an interesting piece of science that breaks new ground. Although it does not detract from the scientific value of their study, the authors have been beguiled by our ripe red herring. Still, they have elegantly streamlined the malodorous query in a way that will allow me to more easily explain the stench, and so I retell their work here. First, I’ll explain some of the new data then I’ll point out where I think they, like many others before them, lost the main scent trail.
The authors examined a box jellyfish, a group that includes the infamous sea wasp, the deadly nemesis of Australian beach-goers. Box jellies are cnidarians, the group including corals, anemones, and hydras, which besides sponges, may be the most distant animal relatives of humans. As such, the biology that is shared between humans and cnidarians likely originated very early in the history of animals, hundreds of millions of years ago. With this logic in mind, Kozmik et al focused on elucidating some of the components of the eyes of one particular box jelly that lives near Puerto Rico, a far less noxious variety than the Australian sea wasp. In particular, they elucidated some photoreceptor genes and some genes that produce a shading pigment in the box jelly’s eyes. Choosing these two particular components was no accident. They were chosen because they represent a common minimum definition of “an eye”. Some scientists argue that minimally, an eye must contain photoreceptor cells to register the presence of photons, and some type of shielding pigment to allow the owner to determine the direction of light. By focusing on these two components, Kozmik and colleagues were streamlining the question of whether eyes evolved more than once, asking whether we could find evidence that photoreceptor plus pigment joined forces multiple separate times, or whether their partnership dates back to a singular union.
Photoreceptor components
Simplifying the real biology a bit, there are two primary classes of phototransduction, which is the cellular process sensing a photon and firing a nervous signal. One pathway that I will call “r” was first elucidated in flies, the other that I will call “c” was first elucidated in vertebrates. It was once thought that the r-pathway is an invertebrate trait, and the c-pathway is a vertebrate trait. However, the last 5 years have brought good evidence that c and r are present in both vertebrates and invertebrates, although the c-pathway is a bit cryptic in invertebrates, and the r-pathway is a bit cryptic in vertebrates. Kozmik et al investigated three components of phototransduction, all three components showed some evidence of being more similar to the “c” class of phototransduction. This primary conclusion led Kozmik et al to choose their title “Assembly of the cnidarian camera-type eye from vertebrate-like components”.
The first component they studied was opsin, a protein that binds to a light reactive chemical to initiate light perception in animals. With opsin, they argued for “c” type phototransduction, as opposed to the “r” type of fly compound eyes. Opsins form multiple sub-families, and the box-jelly opsin might be within the c-opsin subfamily based on their phylogenetic analyses. However, the analysis presented in the supplement of this paper is actually rather inconclusive about c-type status, although it is definitely not an “r”-type. We did a quick reanalysis and found the box jelly opsin might actually belong to a cnidarian-specific opsin sub-family that we discovered last year and which we named “cnidops”. None of the cnidops genes were included in the Kozmik phylogenetic analysis.
Regardless of its sub-family type, this opsin was found to be expressed specifically in the box jelly eyes. One of the most interesting new pieces of science in this paper is that Kozmik et al were able to match the function of the protein itself with that of the jellyfish eye. To do this, they functionally activated box jelly opsin in cell culture and measured the protein’s function. Cell culture is the practice of growing isolated animal cells under controlled conditions, without an animal itself. Introducing the box jelly opsin protein to cells in culture allowed the protein to fold properly and bind to a light- reactive chemical that was separately introduced into the cells. By shining lights of particular wavelengths (“colors”) on to the cells, the authors could see which colors of light activated the box jelly opsin protein. As expected if the opsin in question is used in box jelly eyes, the authors found that the opsin is sensitive to similar wavelengths as the jellyfish eye itself (light we would call blue-green, of about 465 nm wavelength). The match between the animal's light response and that of the expressed opsin protein is a very good piece of evidence supporting the idea the opsin they studied is involved in the animal's light response. This is one of the first times that an invertebrate opsin has been studied in cell culture, and certainly the first Cnidarian opsin, so this is a very exciting experiment.
Evidence for other components of the “c”-pathway being used in box jelly eyes are preliminary. The authors did find that the mRNA of genes similar to vertebrate c-pathway genes were present in the box jelly eye. However, this was as far as the present study went with the non-opsin components. Namely, they did not yet investigate protein expression, nor did they yet conduct any biochemical experiments to demonstrate the interaction of opsin and the other phototransduction components they found. Granted, these additional experiments represent a lot of work. The authors also did not report any experiments to show that r-pathway or other similar genes are not expressed in box jelly eyes (but I note they did not mention the presence of these genes in their library of genes from the box jelly eye, so perhaps they really are absent from the library). In the end, it remains a bit of a leap of faith to conclude that the opsin is actually interacting with the other genes, reported only from mRNA. Nevertheless, the authors have taken some important steps in that direction, so for this essay, we can give the authors the benefit of the doubt that a c-pathway is interestingly being used in the box jellyfish eyes. Even if our hunch is true that it is actually our new “cnidops” class of phototransduction, it doesn’t change the main message here; the herring will remain red.
The second criterion, besides the presence of photoactive cells, required to satisfy a minimal definition of an eye, is the presence of one or more pigmented cells. Here again, Kozmik et al found that pigment-components of a box jelly eye are very similar to pigment-components of vertebrate eyes. Here, the data are quite conclusive. The authors found genes homologous to vertebrate melanin genes expressed in parts of the jellyfish eyes that contain abundant pigment. In addition, there is a direct chemical test for melanin, which box jelly eyes passed. Interestingly, box jelly cells have dual function as photoreceptor and pigment-bearing cell.
The conclusion
Based on these data, Kozmik and colleagues favor the conclusion of parallel evolution – the separate assembly of homologous (“the same”) components to form eyes. In other words, they argue that box jellies and vertebrates separately evolved eyes, but happened to use the same components in each case, opsin and melanin. This conclusion is a bit of a curveball given how I introduced the study above. The conclusion is that the jelly and vertebrate eyes share the same components, so why do they not conclude a common ancestry of the eyes? They actually do point out that this is an alternative possibility. However, they favor parallel evolution based on what boils down to inferring phylogenetic history of these components and of eyes more generally. Unfortunately, they only use verbal arguments, instead of the well established statistics of phylogenetics and ancestral state reconstruction, which could’ve been used to test their claims statistically (see this paper for examples).
The main verbal argument of Kozmik for parallelism, and against common ancestry, is that many animal phyla do not have eyes. As such, it would seem that eyes would have to be lost in many phyla, if common ancestry of box jelly and vertebrate eyes holds (this is where a real phylogenetic statistical test would be nice – undergraduate honor’s project anyone?). In addition, they point out that cnidarians and vertebrates utilize different transcription factor genes in the specification of eyes during development. So basically, they are assuming a priori, based on previous evidence, that the eyes are of “independent origin”, and so based on that assumption, it is surprising to find that they use the same components.
Back to the red herring
So, both vertebrates and box jelly eyes may use c-pathway photoreception and both use melanin as a shielding pigment. You might be wondering where I stand – for parallelism or for common ancestry? Arrrooofff. Bow, wow, wow. Did I catch you? Has your mind been bounding down that pungent scent trail, sniffing and howling enthusiastically? Or have you kept the main, fainter trail, taking my advice that this question of number of origins is misplaced? Perhaps some more explanation will help you understand my point, and wizen your inner bloodhound. Let’s follow the red herring trail and see where it leads.
First, in case it’s not already clear, I’ll point out that the question of parallelism versus common ancestry of box jelly and vertebrate eyes is asking about the number of eye origins. It is our red herring question. Parallelism implies two origins, two separate unions of photoreceptor and pigment. Common ancestry implies a single merger, at least for the two eyes in question.
The reason I don’t like the red herring question, is that it puts the focus on “the eye” rather than on the components that define the eye. The Kozmik et al study is about the components – they elucidated for the first time some of the molecular components of a box jelly eye. This is noteworthy. But what do we gain from saying that eyes originated more than once? I think very little, and in fact such a conclusion masks some of the interesting biology. For example, stating that “eyes originated more than once” fails to recognize the common ancestry of some of the components, notably opsin. Any animal eye that has been examined to date, in addition to photoreceptive cells not in eyes, utilizes a homologous gene family (opsin). This point gets lost when simply stating that eyes evolved more than once.
A related difficulty is that this perspective neglects an intermediate stage in the complexity of photoreceptive structures. Someone unfamiliar (say an anti-evolutionist) who reads “eyes evolved more than once”, imagines the genesis of complex eyes from nothing. But that is not how evolution works. Evolution uses building blocks used for other functions in new combinations. This is illustrated by the conclusion of parallelism of Kozmik et at. In their model, opsin-based phototransduction existed unassociated with pigment cells. Once phototransduction and pigments became associated, perhaps separately in the box jelly and vertebrate lineages, the organisms came to posses a minimum definition of “an eye”. But this definition is just a human construct. Again, to my mind it obscures the fact that much of “the eye” was already there, it just took a new association of components (photoreception plus pigment) to evolve an eye. [For the cynics arguing that this just pushes back the real origins, I recognize this, which is why my lab is studying the origins of phototransduction – by studying the evolutionary history of each of the components separately. The bottom line – phototransduction evolved by using existing and newly duplicated and diverged components, a common theme in evolution].
You may be asking what if the opposite answer of the red herring question holds. What if we conclude “eyes share a common ancestry”, which could be concluded based on the Kozmik et al data of shared opsin and pigment components in vertebrates and box jellies. Again, I would argue that we gain little by making this statement. In this case, such a statement can only be made for the components in question. Perhaps it can serve as a sort of “null hypothesis”, where it serves as a prediction for new data that we don’t yet know about, which has some value. However, we already know that for eyes, not all components have the same history, which is implied by the statement “eyes only evolved once”. We already know that different lens proteins are used in different types of eyes. We also already know that components of phototransduction cascades differ. For example c-phototransduction utilizes the ion channel protein CNG, whereas r-pathways utilize the unrelated ion channel protein TRP. Saying that all eyes have a common ancestry does not account for these details. Again, it is about the components. Different components have different histories, and this cannot be accounted for in the simple dichotomy of “one versus more than one origin”.
The question of the number of eye origins has been around for a while now. But I argue that it is time to move beyond this question, and to recognize it for the red herring that it is. The fact of the matter is that some eye components are shared and others are not, and this varies depending on the time scale and species examined. Of value is to fill in the narrative of this amazing evolutionary story. We now have the ability to understand to the level of specific molecular changes how vastly different types of eyes evolved. This is the main – though perhaps fainter – scent trail.
I've come to realize that my own field of eye evolution has a resoundingly pungent red herring that continually throws scientists and non-scientists alike off the trail of a deep understanding of how eyes, and more generally, photoreception, evolved. In this case, the rank fish is the simple and seemingly harmless question, "How many times did eyes evolve?"
Quite often, when I tell a newly acquainted colleague that I study eye evolution, he or she will pull the red herring out of their back pocket, and dash across the room, dragging the fish behind them. "So how many times did eyes evolve, anyway?” they ask, hoping I have a key to this deep mystery. When I was a younger bloodhound, I often took that bait, howling and bounding enthusiastically after that trail, circling the room again and again. But I've followed that trail enough times by now to know that it circles back on itself in a confusing and illogical way.
How many times eyes evolved, though enticingly simple, is an all but unanswerable question; at least in terms of a numerical answer that people crave. Eyes did not evolve one time, nor did they evolve forty to sixty-five times. The answer is that some components of animal eyes are shared in all animal eyes we've ever examined. Yet some components are new comers to organs that probably functioned as eyes even before those components joined the party. Asking how many times eyes evolved makes the implicit assumption that every component of that eye shares a congruent history, that an eye is either all there, or all not there. But eyes are complicated structures, and the evolutionary histories of its parts are varied.
A new paper by Kozmik et al in PNAS is an interesting piece of science that breaks new ground. Although it does not detract from the scientific value of their study, the authors have been beguiled by our ripe red herring. Still, they have elegantly streamlined the malodorous query in a way that will allow me to more easily explain the stench, and so I retell their work here. First, I’ll explain some of the new data then I’ll point out where I think they, like many others before them, lost the main scent trail.
The authors examined a box jellyfish, a group that includes the infamous sea wasp, the deadly nemesis of Australian beach-goers. Box jellies are cnidarians, the group including corals, anemones, and hydras, which besides sponges, may be the most distant animal relatives of humans. As such, the biology that is shared between humans and cnidarians likely originated very early in the history of animals, hundreds of millions of years ago. With this logic in mind, Kozmik et al focused on elucidating some of the components of the eyes of one particular box jelly that lives near Puerto Rico, a far less noxious variety than the Australian sea wasp. In particular, they elucidated some photoreceptor genes and some genes that produce a shading pigment in the box jelly’s eyes. Choosing these two particular components was no accident. They were chosen because they represent a common minimum definition of “an eye”. Some scientists argue that minimally, an eye must contain photoreceptor cells to register the presence of photons, and some type of shielding pigment to allow the owner to determine the direction of light. By focusing on these two components, Kozmik and colleagues were streamlining the question of whether eyes evolved more than once, asking whether we could find evidence that photoreceptor plus pigment joined forces multiple separate times, or whether their partnership dates back to a singular union.
Photoreceptor components
Simplifying the real biology a bit, there are two primary classes of phototransduction, which is the cellular process sensing a photon and firing a nervous signal. One pathway that I will call “r” was first elucidated in flies, the other that I will call “c” was first elucidated in vertebrates. It was once thought that the r-pathway is an invertebrate trait, and the c-pathway is a vertebrate trait. However, the last 5 years have brought good evidence that c and r are present in both vertebrates and invertebrates, although the c-pathway is a bit cryptic in invertebrates, and the r-pathway is a bit cryptic in vertebrates. Kozmik et al investigated three components of phototransduction, all three components showed some evidence of being more similar to the “c” class of phototransduction. This primary conclusion led Kozmik et al to choose their title “Assembly of the cnidarian camera-type eye from vertebrate-like components”.
The first component they studied was opsin, a protein that binds to a light reactive chemical to initiate light perception in animals. With opsin, they argued for “c” type phototransduction, as opposed to the “r” type of fly compound eyes. Opsins form multiple sub-families, and the box-jelly opsin might be within the c-opsin subfamily based on their phylogenetic analyses. However, the analysis presented in the supplement of this paper is actually rather inconclusive about c-type status, although it is definitely not an “r”-type. We did a quick reanalysis and found the box jelly opsin might actually belong to a cnidarian-specific opsin sub-family that we discovered last year and which we named “cnidops”. None of the cnidops genes were included in the Kozmik phylogenetic analysis.
Regardless of its sub-family type, this opsin was found to be expressed specifically in the box jelly eyes. One of the most interesting new pieces of science in this paper is that Kozmik et al were able to match the function of the protein itself with that of the jellyfish eye. To do this, they functionally activated box jelly opsin in cell culture and measured the protein’s function. Cell culture is the practice of growing isolated animal cells under controlled conditions, without an animal itself. Introducing the box jelly opsin protein to cells in culture allowed the protein to fold properly and bind to a light- reactive chemical that was separately introduced into the cells. By shining lights of particular wavelengths (“colors”) on to the cells, the authors could see which colors of light activated the box jelly opsin protein. As expected if the opsin in question is used in box jelly eyes, the authors found that the opsin is sensitive to similar wavelengths as the jellyfish eye itself (light we would call blue-green, of about 465 nm wavelength). The match between the animal's light response and that of the expressed opsin protein is a very good piece of evidence supporting the idea the opsin they studied is involved in the animal's light response. This is one of the first times that an invertebrate opsin has been studied in cell culture, and certainly the first Cnidarian opsin, so this is a very exciting experiment.
Evidence for other components of the “c”-pathway being used in box jelly eyes are preliminary. The authors did find that the mRNA of genes similar to vertebrate c-pathway genes were present in the box jelly eye. However, this was as far as the present study went with the non-opsin components. Namely, they did not yet investigate protein expression, nor did they yet conduct any biochemical experiments to demonstrate the interaction of opsin and the other phototransduction components they found. Granted, these additional experiments represent a lot of work. The authors also did not report any experiments to show that r-pathway or other similar genes are not expressed in box jelly eyes (but I note they did not mention the presence of these genes in their library of genes from the box jelly eye, so perhaps they really are absent from the library). In the end, it remains a bit of a leap of faith to conclude that the opsin is actually interacting with the other genes, reported only from mRNA. Nevertheless, the authors have taken some important steps in that direction, so for this essay, we can give the authors the benefit of the doubt that a c-pathway is interestingly being used in the box jellyfish eyes. Even if our hunch is true that it is actually our new “cnidops” class of phototransduction, it doesn’t change the main message here; the herring will remain red.
The second criterion, besides the presence of photoactive cells, required to satisfy a minimal definition of an eye, is the presence of one or more pigmented cells. Here again, Kozmik et al found that pigment-components of a box jelly eye are very similar to pigment-components of vertebrate eyes. Here, the data are quite conclusive. The authors found genes homologous to vertebrate melanin genes expressed in parts of the jellyfish eyes that contain abundant pigment. In addition, there is a direct chemical test for melanin, which box jelly eyes passed. Interestingly, box jelly cells have dual function as photoreceptor and pigment-bearing cell.
The conclusion
Based on these data, Kozmik and colleagues favor the conclusion of parallel evolution – the separate assembly of homologous (“the same”) components to form eyes. In other words, they argue that box jellies and vertebrates separately evolved eyes, but happened to use the same components in each case, opsin and melanin. This conclusion is a bit of a curveball given how I introduced the study above. The conclusion is that the jelly and vertebrate eyes share the same components, so why do they not conclude a common ancestry of the eyes? They actually do point out that this is an alternative possibility. However, they favor parallel evolution based on what boils down to inferring phylogenetic history of these components and of eyes more generally. Unfortunately, they only use verbal arguments, instead of the well established statistics of phylogenetics and ancestral state reconstruction, which could’ve been used to test their claims statistically (see this paper for examples).
The main verbal argument of Kozmik for parallelism, and against common ancestry, is that many animal phyla do not have eyes. As such, it would seem that eyes would have to be lost in many phyla, if common ancestry of box jelly and vertebrate eyes holds (this is where a real phylogenetic statistical test would be nice – undergraduate honor’s project anyone?). In addition, they point out that cnidarians and vertebrates utilize different transcription factor genes in the specification of eyes during development. So basically, they are assuming a priori, based on previous evidence, that the eyes are of “independent origin”, and so based on that assumption, it is surprising to find that they use the same components.
Back to the red herring
So, both vertebrates and box jelly eyes may use c-pathway photoreception and both use melanin as a shielding pigment. You might be wondering where I stand – for parallelism or for common ancestry? Arrrooofff. Bow, wow, wow. Did I catch you? Has your mind been bounding down that pungent scent trail, sniffing and howling enthusiastically? Or have you kept the main, fainter trail, taking my advice that this question of number of origins is misplaced? Perhaps some more explanation will help you understand my point, and wizen your inner bloodhound. Let’s follow the red herring trail and see where it leads.
First, in case it’s not already clear, I’ll point out that the question of parallelism versus common ancestry of box jelly and vertebrate eyes is asking about the number of eye origins. It is our red herring question. Parallelism implies two origins, two separate unions of photoreceptor and pigment. Common ancestry implies a single merger, at least for the two eyes in question.
The reason I don’t like the red herring question, is that it puts the focus on “the eye” rather than on the components that define the eye. The Kozmik et al study is about the components – they elucidated for the first time some of the molecular components of a box jelly eye. This is noteworthy. But what do we gain from saying that eyes originated more than once? I think very little, and in fact such a conclusion masks some of the interesting biology. For example, stating that “eyes originated more than once” fails to recognize the common ancestry of some of the components, notably opsin. Any animal eye that has been examined to date, in addition to photoreceptive cells not in eyes, utilizes a homologous gene family (opsin). This point gets lost when simply stating that eyes evolved more than once.
A related difficulty is that this perspective neglects an intermediate stage in the complexity of photoreceptive structures. Someone unfamiliar (say an anti-evolutionist) who reads “eyes evolved more than once”, imagines the genesis of complex eyes from nothing. But that is not how evolution works. Evolution uses building blocks used for other functions in new combinations. This is illustrated by the conclusion of parallelism of Kozmik et at. In their model, opsin-based phototransduction existed unassociated with pigment cells. Once phototransduction and pigments became associated, perhaps separately in the box jelly and vertebrate lineages, the organisms came to posses a minimum definition of “an eye”. But this definition is just a human construct. Again, to my mind it obscures the fact that much of “the eye” was already there, it just took a new association of components (photoreception plus pigment) to evolve an eye. [For the cynics arguing that this just pushes back the real origins, I recognize this, which is why my lab is studying the origins of phototransduction – by studying the evolutionary history of each of the components separately. The bottom line – phototransduction evolved by using existing and newly duplicated and diverged components, a common theme in evolution].
You may be asking what if the opposite answer of the red herring question holds. What if we conclude “eyes share a common ancestry”, which could be concluded based on the Kozmik et al data of shared opsin and pigment components in vertebrates and box jellies. Again, I would argue that we gain little by making this statement. In this case, such a statement can only be made for the components in question. Perhaps it can serve as a sort of “null hypothesis”, where it serves as a prediction for new data that we don’t yet know about, which has some value. However, we already know that for eyes, not all components have the same history, which is implied by the statement “eyes only evolved once”. We already know that different lens proteins are used in different types of eyes. We also already know that components of phototransduction cascades differ. For example c-phototransduction utilizes the ion channel protein CNG, whereas r-pathways utilize the unrelated ion channel protein TRP. Saying that all eyes have a common ancestry does not account for these details. Again, it is about the components. Different components have different histories, and this cannot be accounted for in the simple dichotomy of “one versus more than one origin”.
The question of the number of eye origins has been around for a while now. But I argue that it is time to move beyond this question, and to recognize it for the red herring that it is. The fact of the matter is that some eye components are shared and others are not, and this varies depending on the time scale and species examined. Of value is to fill in the narrative of this amazing evolutionary story. We now have the ability to understand to the level of specific molecular changes how vastly different types of eyes evolved. This is the main – though perhaps fainter – scent trail.
Subscribe to:
Posts (Atom)